Бластула, называемая также зародышевым пузырем, является окончательным результатом процесса дробления яйца. Следующая за ним стадия, занимающая промежуточное положение между дроблением и органогенезом, в эмбриогенезе это гаструляция. Ее основной смысл заключается в образовании трех зародышевых листков: энтодермы, эктодермы и мезодермы. Иными словами, именно с гаструляции начинается эмбриональное дифференцирование и морфогенез организма.
Определение термина "гаструляция"
Еще в далеком 1901 году гаструляция была описана как путь, по которому внутрь эмбриона попадают мезодермальные, энтодермальные и эктодермальные клетки. Данное определение подразумевает наличие у бластулы особых органообразующих пространств. Поняв это довольно простое описание, легко перейти к более сложному, современному значению термина. Гаструляция - это последовательность морфогенетических движений, результатом которых является перемещение зачатков тканей в места, предназначенные для них в соответствии с «планом» организации организма. Процесс сложный, изменения сопровождаются ростом и размножением, направленным перемещением и дифференцированием клеток.
Рассматривая гаструляцию в более общем смысле, можно определить ее как промежуточный этап, принадлежащий к единому динамическому процессу, в течение которого происходит перестройка участков бластулы, что заметно облегчает переход к процессу органогенеза.
Перемещение клеток
Если давать общую характеристику рассматриваемому процессу, то можно сказать, что гаструляция - это эмболия и эпиболия. Оба термина отражают морфогенетическое движение клеток, которое происходит абсолютно на всех этапах онтогенетического развития организма. Однако наиболее выражены они именно при гаструляции. Эпиболией называют процесс перемещения клеток по поверхности зародыша, а эмболией - их движение внутрь него.
В эмбриологии различают следующие основные типы гаструляции или движения клеток: инвагинация, иммиграция, инволюция, деламинация и эпиболия. Более подробно о них - далее в статье.
Перемещение клеточных пластов
Принимать участие в процессе гаструляции могут не только отдельные (свободно мигрирующие) клетки, но и целые клеточные пласты. Направление определяют константное и дистантное взаимодействия. Первые силы были открыты П. Вейсом в 20-х годах прошлого столетия и случаются, по-видимому, и в эмбриогенезе, вторые - редкие и специальные, при обычном морфогенезе происходят с малой долей вероятности.
При гаструляции дробление клетки не происходит. Как уже было сказано выше, начинается движение клеточных масс и в результате образование двухслойного зародыша, называемого гаструлой. Становятся четко заметны энтодерма и эктодерма. У всех многоклеточных организмов (исключением являются лишь кишечнополостные) параллельно с гаструляцией либо сразу после нее формируется третий зародышевый листок, именуемый мезодермой. Он является совокупностью клеток, расположенных между эктодермой и энтодермой. В результате зародыш становится трехслойным.
Способы гаструляции напрямую зависят от типа бластулы.
Инвагинационная гаструла

Название способа говорит само за себя. Инвагинация - это впячивание однослойной стенки бластулы (балстодермы) внутрь бластоцеля. Наиболее примитивным и максимально наглядным будет пример с резиновым мячиком. При нажатии на него часть материала вдавливается вовнутрь. Впячивание можно довести до самой дальней стенки либо сделать его незначительным. В результате происходит преобразование бластулы, и гаструла получается в виде двухслойного мешка с архентероном. Ее внутренней стенкой является первичная энтодерма, а наружной - первичная эктодерма. Образовавшийся при этом архентерон (первичный кишечник) сообщается с наружной средой при помощи отверстия, называемого бластопором. Второе его название - первичный рот. Дальнейшее его развитие зависит от вида организма. У многих животных бластопор в конечном итоге преобразуется в дефинитивный рот. В связи с этим их называют первичноротыми (моллюски, черви, членистоногие). У вторичноротых бластопор превращается в нервно-кишечный канал, расположенный в задней части эмбриона (у хордовых), либо в анальное отверстие.
Иммиграционная гаструла
Иммиграционная гаструляция - это способ образования двухслойного эмбриона, наиболее характерный для кишечнополостных. Гаструла образуется путем активного выселения внутрь бластоцеля части клеток бластулы. Такая иммиграция носит униполярный характер. Клетки перемещаются лишь с вегетативного полюса. Позднее они образуют энтодерму, т. е. внутренний слой. Именно таким способом гаструляция осуществляется у гидроидного полипа, медузы.
Клетки бластодермы в бластоцель могут проникать ни в каком-либо одном участке, а по всей поверхности зародыша. Такая иммиграция получила название мультиполярной, но встречается она довольно редко.
У многих кишечнополостных, которым свойственен иммиграционный способ гаструляции, происходит очень активное «выселение» клеток бластулы, и гаструла, образовавшаяся в итоге, полностью теряет бластоцель. В данном случае характерный для предыдущего инвагинационного способа бластопор отсутствует.
Деламинационная гаструла

Данный редкий вид гаструлы впервые был описан Мечниковым И. И., и характерен он для кишечнополостных. Сопровождающие гаструляцию процессы весьма своеобразны, но при рассмотрении типичного случая они воспринимаются проще. Например, яйца некоторых сцифомедуз обладают концентрически расположенными и хорошо различимыми участками цитоплазмы: плотная и зернистая (эктоплазма) и ячеистая (эндоплазма). Для них характерно относительно синхронное и равномерное деление: 2, 4, 8, 16. В конечном итоге зародыш содержит 32 бластомера. Далее деление осуществляется параллельно поверхности зародыша. Образуется наружный слой бластомеров, состоящий из эктоплазмы, и внутренний - частично из эктоплазмы и из эндоплазмы. Иначе говоря, процесс образования многослойного зародыша идет путем расщепления одного слоя клеток на два. Затем дробятся только внутренние бластомеры и вновь параллельно поверхности зародыша, который в результате столь своеобразной гаструляции обретает форму шара. Он состоит из 64 образующих эктодерму плоских клеток и 32 более выпуклых, являющихся основой энтодермы.
Эпиболическая гаструла

У животных, имеющих резко выраженное телолецитальное строение яиц (смещение желтка к вегетативному полюсу), гаструляция происходит по эпиболическому способу. Макромеры представляют собой крупные бластомеры, которые делятся очень медленно и содержат большое количество желтка. У них отсутствует способность к перемещению, в связи с этим на них буквально «наползают» более активные микромеры, расположенные на поверхности клетки. При такой гаструляции бластопор отсутствует, а архентерон не образуется. Лишь в дальнейшем, когда макромеры все же уменьшаются в размерах, начинает формироваться полость, зачаток первичного кишечника.
Инволюция
Инволюционная гаструляция - это процесс, заключающийся в «подворачивании» наружного пласта клеток вовнутрь зародыша. Он, увеличиваясь в размерах, распространяется по внутренней поверхности. Данный способ гаструляции характерен для животных с мезолецитальными яйцеклетками - амфибий (земноводных). Перемещение лидирующих глубоких клеток краевой области тормозит развитие архентерона. Именно в них заключена движущая сила инволюции.

Смешанный способ гаструляции
Как известно, эмбриогенез - это самый ранний период развития каждого индивидуального организма: от зачатия до рождения. Гаструляция является одним из его этапов, вторым по хронологии после дробления. Ее способы настолько различны, что сравнивать их можно с большой долей условности. Каждый из них требует детального изучения и анализа. Однако есть все же определенные линии пересечения между ними. Так, в качестве своеобразного варианта инвагинации можно рассматривать процесс эпиболии, а деламинация имеет черты сходства с иммиграцией.
Отметим, что у многих животных гаструляция идет комбинированным способом. В таких случаях одновременно имеют место эпиболия и инвагинация, а также иные морфогенетические процессы. В частности, именно так гаструляция протекает у амфибий. В связи с этим многие авторы выделяют смешанный способ.
Гаструла

Дословно с латыни термин «гаструла» переводится как «чрево, желудок». Он обозначает специфическую зародыша многоклеточных организмов. Отличительная особенность гаструлы - это наличие двух или трех зародышевых листков. Процесс ее образования - это фаза гаструляции.
Наиболее простое устройство наблюдается у животных. Для них характерна гаструла эллипсоидной формы с одноклеточным внешним слоем (эктодерма) и внутренним скоплением клеток (энтодерма), а также «первичной кишкой». Типичной считают гаструлу морского ежа, которая образуется путем инвагинации. У человека гаструляция проходит на 8-9-е сутки развития. Гаструла представляет собой дисковидное уплощенное образование, сформированное из внутренней клеточной массы.
Как правило, у большинства животных на стадии гаструлы зародыш свободно жить не может и располагается в матке или яйцевых оболочках. Однако существуют и исключения. Так, личинки кишечнополостных, планулы, представляют собой свободноплавающую гаструлу.
ЭМБРИОЛОГИЯ
Занятие №4
ТЕМА: «Эмбриогенез. Дробление, гаструляция»
КОНТРОЛЬНЫЕ ВОПРОСЫ
Определение понятия «дробление».
Характер дробления зиготы у человека.
Темные и светлые бластомеры, их морфологическая характеристика, потенции развития.
Морула и бластоциста, их морфологическая характеристика.
Имплантация зародыша в слизистую оболочку матки. Стадии имплантации.
Гистиотрофный и гематотрофный типы питания зародыша.
Способы гаструляции у человека.
Стадии гаструляции (деламинация и иммиграция), их сроки и механизмы.
Факторы, влияющие на механизмы гаструляции.
ЦЕЛЬ ЗАНЯТИЯ:
Усвоить особенности и характер дробления зиготы у человека. Научиться различать темные и светлые бластомеры. Усвоить различие между морулой и бластоцистой. Знать стадии имплантации и типы питания зародыша в зависимости от стадии имплантации. Изучить способы гаструляции у человека, знать стадии и механизмы гаструляции.
Студент должен знать:
характер дробления зиготы у человека;
морфологическую характеристику темных и светлых бластомеров, потенции развития;
морфологическую характеристику морулы и бластоцисты;
стадии имплантации зародыша в слизистую оболочку матки;
способы гаструляции у человека, их сроки и механизмы.
Студент должен уметь:
на схемах и таблицах уметь различать темные и светлые бластомеры; дифференцировать морулу и бластоцисту;
различать стадии имплантации зародыша в слизистую оболочку матки;
различать способы гаструляции у человека;
сделать зарисовки и составить их письменный протокол.
ДРОБЛЕНИЕ
Дробление представляет собой серию митотических делений зиготы с образованием многих дочерних клеток (бластомеров) меньшего размера. Митотические деления зиготы и в последующем бластомеров происходят с увеличением числа клеток, но без увеличения их массы, поэтому именуются дроблением.
Типы дробления:
полное, или голобластическое, (ланцетник, амфибии, млекопитающие): зигота полностью разделяется на бластомеры;
частичное, или меробластическое, (рыбы, рептилии, птицы): дробится лишь часть зиготы;
равномерное: образующиеся бластомеры одинаковы или близки по величине;
неравномерное: бластомеры отличаются по размерам;
синхронное: бластомеры делятся одновременно;
асинхронное.
По характеру пространственного расположения бластомеров выделяют следующие виды полного дробления :
радиальное: образующиеся бластомеры располагаются друг над другом, создавая фигуру с радиальной симметрией (ланцетник, амфибии);
спиральное: вышележащие бластомеры смещены по отношению к нижележащим под углом 45°, располагаясь, таким образом, по спирали (моллюски, немертины, кольчатые черви и некоторые планарии);
билатеральное (билатерально-симметричное): на ранних стадиях расположение бластомеров происходит по закону билатеральной симметрии, так что бластомерам правой стороны зародыша соответствуют точно такие же бластомеры левой стороны (асцидии, нематоды и некоторые другие беспозвоночные)
хаотическое (неупорядоченное): уже после третьего деления отсутствует строгая закономерность в расположении бластомеров, в ходе дальнейшего дробления возникающая беспорядочность усиливается (некоторые медузы). Внешняя хаотичность, однако, регулируется внутренними механизмами, которые еще не выяснены, и приводит к образованию животного конкретного вида.
Частичное дробление существует в двух видах:
дискоидальное: дробление, при котором на бластомеры делится лишь часть цитоплазмы у анимального полюса (рыбы, рептилии, птицы);
поверхностное: дробится поверхностный слой цитоплазмы, по мере развития полностью обособляющийся от желтка (насекомые и большая часть других членистоногих).
В процессе дробления зародыш перемещается по маточной трубе и заканчивает дробление в полости матки на 6-е сутки развития. Дробление завершается образованием бластулы. Это многоклеточный зародыш, как правило, с полостью внутри. Стенку бластулы называют бластодерма , а полость - бластоцель (первичная полость тела). У бластулы также обычно различают крышу, дно и разграничивающую их краевую зону . Бластулу, образующуюся при полном равномерном дроблении, называют равномерная целобластула (ланцетник), при полном неравномерном - неравномерная целобластула, называемая также амфибластулой (амфибии) или бластоцистой (млекопитающие, человек), при частичном дискоидальном - дискобластула (рептилии, птицы), при поверхностном - стерробластула (кишечнополостные).
На стадии бластулы окончательно устанавливается полярность зародыша, повышается степень его интеграции и возникает именно тот порядок расположения клеток и та степень их взаимодействия, которые необходимы для направленных клеточных движений на следующем этапе развития - гаструляции.
На стадии поздней бластулы у ланцетника и низших позвоночных устанавливаются так называемые презумптивные области, которые содержат материал определенных органов и систем. Расположение и границы презумптивных областей, или презумптивных зачатков, были изучены с помощью метода маркировки участков зародыша витальными красителями (метод В. Фогта, предложен в 1925 г.). После нанесения метки на поверхность зародыша можно проследить ее перемещения и выяснить судьбу данного участка в ходе гаструляции и дальнейшего развития.
План взаимного расположения презумптивных зачатков будущих органов получил название карты презумптивных зачатков.Карта зачатков, например, у амфибий, точно указывает положение областей, из которых будут развиваться эпидермис кожи, органы обоняния, слуха, хрусталик глаза и другие.
Дробление зародыша человека полное, или голобластическое (борозды дробления проходят через весь зародыш), неравномер ное (в результате дробления образуются бластомеры неравной величины)и асинхронное (разные бластомеры дробятся с различной скоростью, поэтому зародыш на отдельных стадиях дробления содержит нечетное число клеток). Первое дробление (деление) зиготы завершается через 30 ч, в результате образуются два бластомера, покрытых оболочкой оплодотворения, За стадией двух бластомеров следует стадия трех бластомеров.
С первых же дроблений зиготы формируются два вида бластомеров - «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и внезародышевые органы (амнион, желточный мешок, аллантоис).
Начиная с 3 суток, дробление идет быстрее, и на 4-е сутки зародыш состоит из 7-12 бластомеров. Через 50 - 60 ч образуется плотное скопление клеток - морула , а на 3-4-е сут. начинается формирование бластоцисты - полого пузырька, заполненного жидкостью. Бластоциста в течение 3 суток перемещается по маточной трубе к матке и через 4 суток попадает в полость матки. Бластоциста находится в полости матки в свободном виде (свободная бластоциста) в течение 2 суток (5-е и 6-е сутки). К этому времени бластоциста увеличивается в размере благодаря росту числа бластомеров - клеток эмбриобласта и трофобласта - до 100 и вследствие усиленного всасывания трофобластом секрета маточных желез и активной выработки жидкости клетками трофобласта.
Трофобласт первые 2 недели развития обеспечивает питание зародыша за счет продуктов распада материнских тканей – гистиотрофный тип питания. Эмбриобласт располагается в виде узелка зародышевых клеток («зародышевый узелок»), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты.
ИМПЛАНТАЦИЯ
Имплантация (врастание, укоренение) - внедрение зародыша в слизистую оболочку матки. Различают две стадии имплантации:
адгезию (прилипание): зародыш прикрепляется к внутренней поверхности матки;
инвазию (погружение) - внедрение зародыша в ткани слизистой оболочки матки.
На 7-е сут. в трофобласте и эмбриобласте происходят изменения, связанные с подготовкой к имплантации:
бластоциста сохраняет оболочку оплодотворения;
в трофобласте увеличивается количество лизосом с ферментами, обеспечивающими разрушение (лизис) тканей стенки матки и тем самым способствующими внедрению зародыша в толщу ее слизистой оболочки;
появляющиеся в трофобласте микроворсинки постепенно разрушают оболочку оплодотворения;
зародышевый узелок уплощается и превращается в зародышевый щиток, в котором начинается подготовка к первой стадии гаструляции.
Имплантация продолжается около 40 часов. Одновременно с имплантацией начинается гаструляция (образование зародышевых листков). Это первый критический период развития.
В первой стадии трофобласт прикрепляется к эпителию слизистой оболочки матки, и в нем формируются два слоя - цитотрофобласт и симпластотрофобласт.
Во второй стадии симпластотрофобласт, продуцируя протеолитические ферменты, разрушает слизистую оболочку матки. Формирующиеся при этом ворсинки трофобласта, внедряясь в матку, последовательно разрушают ее эпителий, затем подлежащую соединительную ткань и стенки сосудов, и трофобласт вступает в непосредственный контакт с кровью материнских сосудов. Питание зародыша осуществляется непосредственно из материнской крови (гематотрофный тип питания). Из крови матери зародыш получает не только все питательные вещества, но и кислород, необходимый для дыхания.
Гематотрофный тип питания, сменяющий гистиотрофный, сопровождается переходом к качественно новому этапу эмбриогенеза - второй фазе гаструляции и закладке внезародышевых органов .
Гаструляция
Период гаструляции характеризуется активными перемещениями, как отдельных клеток зародыша, так и клеточных масс, в результате которых у позвоночных формируются три основных пласта тела, их называют зародышевыми листками :
эктодерма - наружный зародышевый листок;
мезодерма - средний зародышевый листок;
энтодерма - внутренний зародышевый листок.
Зародышевые листки у разных животных являются гомологичными образованиями , т.е. в ходе развития они дают идентичные структуры : эктодерма всегда преобразуется в наружный покров тела, а из энтодермы развивается выстилка средней кишки.
Другая особенность периода гаструляции заключается в том, что образующиеся при делении клетки , в отличие от бластомеров, начинают расти и увеличиваются до размеров материнской, при этом происходит активный рост и увеличение размеров самого зародыша.
Гаструляция у человека протекает в две стадии:
I стадия (деламинация) приходится на 7-е сутки внутриутробного развития: из материала зародышевого узелка (эмбриобласта) образуются два листка: наружный листок - эпибласт (клетки имеют вид псевдомногослойного призматического эпителия) и внутренний - гипобласт , обращенный в полость бластоцисты (клетки мелкие кубические, с пенистой цитоплазмой).
II стадия (иммиграция) - на 14-15-е сутки внутриутробного развития.
Способы гаструляции , т. е. механизмы образования зародышевых листков, отличаются у разных животных и определяются во многом строением бластулы. Выделяют четыре основных способа гаструляции:
инвагинация (впячивание);
деламинация (расслоение, расщепление);
иммиграция (выселение);
эпиболия (обрастание).
В результате гаструляции возникает зародыш - гаструла . Гаструла имеет полость - гастроцель (полость первичной кишки), в которую ведет отверстие - бластопор (первичный рот).
В зависимости от дальнейшей судьбы бластопора в развитии все животные подразделяются на первичноротых и вторичноротых. У первичноротых, к которым относится большинство беспозвоночных, на месте бластопора образуется ротовое отверстие, а противоположный конец становится задним концом тела. У вторичноротых, которые включают хордовых и некоторых беспозвоночных, бластопор преобразуется в анальное отверстие или нервнокишечный канал, расположенный на заднем конце тела, а ротовое отверстие прорывается на брюшной стороне у противоположного конца тела. У бластопора различают края, или губы: дорсальную, латеральные и вентральную .
Важным итогом гаструляции у хордовых является формирование в составе зародышевых листков так называемого осевого комплекса зачатков. Осевой комплекс зачатков представляет собой расположенные по оси тела зародыша зачатки нервной системы (нервная пластинка) и хорды (хордальная пластинка), а также лежащие латерально по отношению к хордальной пластинке и связанные с ней зачатки мезодермы. Тесно прилежащие друг к другу зачатки хорды и мезодермы часто называют хордомезо дермой .
Существует несколько факторов гаструляции:
физические: к таковым относится более быстрое деление мелких клеток по сравнению с более крупными, что приводит к обрастанию крупных клеток мелкими, например, в бластуле амфибий и рыб, имеющих телолецитальные яйцеклетки. В бластуле этих животных существует градиент распределения бластомеров по величине - их размеры уменьшаются по направлению от вегетативного полюса к анимальному. Такое же направление имеет метаболический градиент - с уменьшением размеров клеток повышается их метаболическая активность и темпы деления. Именно в областях активного деления клеток начинаются процессы их перемещения. Одной из причин перемещения клеток считают изменение поверхностного натяжения при увеличении числа клеток. Клетки при этом способны совершать активные амебоидные движения;
химические: механизм индуцирующего влияния связан с выделением особых факторов: белков, нуклеопротеидов, стероидов.
организующие центры: направление движения клеток, а затем и их дифференцировка определяются индукцией - влиянием одних областей или зачатков зародыша на другие. Такие области и зачатки получили название организующих центров, или индукторов. Теория организующих центров, выдвинутая Г.Шпеманом, устанавливает наличие у зародыша на разных стадиях его развития особых областей, оказывающих индуцирующее влияние на соседние участки. К организующим центрам, например, относится передняя (дорсальная) губа бластопора, индуцирующая формирование хордомезодермы, которая, в свою очередь, вызывает дифференцировку части клеток эктодермы в нервную пластинку.
Важную роль в регуляции межклеточных взаимодействий во время развития выполняют специализированные межклеточные соединения - щелевые контакты , появляющиеся уже во время дробления между бластомерами и, возможно, представляющие собой первую систему передачи сигналов между клетками.
Типы дробления (ознакомиться)
А - радиальное дробление (ланцетник);
Б - билатеральное дробление (аскарида);
В - неупорядоченное дробление (сосальщики);
Г - спиральное дробление (моллюск).
Цифрами обозначена последовательность стадий дробления.
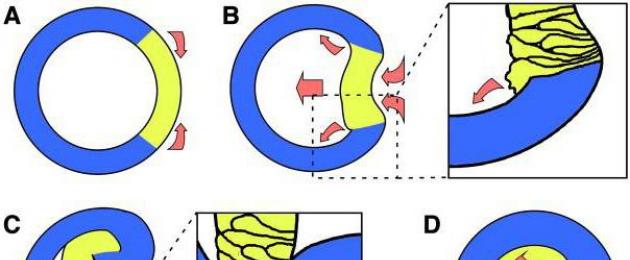
По окончании периода дробления у многоклеточных животных начинается период образования зародышевых листков – гаструляция . Она связана с перемещением эмбрионального материала. Сначала образуется ранняя гаструла, имеющая 2 зародышевых листка (эктодерму и энтодерму), затем поздняя гаструла, когда формируется третий зародышевый листок – мезодерма. Образующийся зародыш называют гаструлой.
Образование ранней гаструлы может происходить несколькими способами: иммиграцией, инвагинацией, эпиболией или деляминацией (рис. 5).
При иммиграции (выселении) часть клеток бластодермы с поверхности зародыша уходит в бластоцель. Образуется наружный слой (эктодерма) и внутренний (энтодерма). Бластоцель заполнена клетками. Такой способ образования гаструлы характерен, например, для кишечнополостных.
Инвагинация (впячивание) наблюдается в случае целобластулы. При инвагинации определенный участок бластодермы (вегетативный полюс) прогибается внутрь и достигает анимального полюса. Образуется двуслойный зародыш – гаструла. Наружный слой клеток называют эктодермой, внутренний – энтодермой. Энтодерма выстилает полость первичной кишки – гастроцель. Отверстие, при помощи которого полость сообщается с внешней средой, называется первичным ртом – бластопором. По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми (хордовые).
Эпиболия (обрастание) характерна для животных, развивающихся из телолецитальных яиц. Образование гаструлы идет за счет быстрого деления микромеров, которыми обрастает вегетативный полюс.
Рис. 5. Типы гаструляции (Ю.П. Антипчук, 1983)
I – инвагинация; II – эпиболия, III – иммиграция, IV – деляминация.
Макромеры оказываются внутри зародыша. Образования бластопора не происходит, нет гастроцели. Такой способ гаструляции наблюдается у круглоротых и земноводных.
Деляминация (расслоение) встречается у организмов, бластула которых похожа на морулу. Клетки бластодермы делятся на наружный и внутренний слои. Наружный слой образует эктодерму, внутренний – энтодерму. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных.
У человека гаструляция протекает в две фазы. Первая фаза (7-е сутки) происходит путем деляминации эмбриобласта. Образуются два листка: наружный - эпибласт и внутренний - гипобласт. Вторая стадия (14-15 сутки) происходит с образованием первичной полоски и первичного узелка путем перемещения, иммиграции клеточных масс.
У всех многоклеточных, кроме губок и кишечнополостных, образуется третий зародышевой листок – мезодерма . Он может формироваться четырьмя способами (рис. 6).
Телобластический - мезодерма образуется за счет нескольких крупных клеток на заднем конце зародыша - телобластов, которые располагаются между эктодермой и энтодермой. За счет расслоения клеток мезодермы образуется вторичная полость тела - целом. Такой способ образования мезодермы характерен для первичноротых животных.
Энтероцельный - мезодерма образуется из клеток энтодермы одновременно с формированием целома. Характерно для вторичноротых животных.
Эктодермальный - мезодерма образуется из части клеток эктодермы, которые размещаются между ней и энтодермой. Такой способ образования мезодермы характерен для пресмыкающихся, птиц, млекопитающих и человека.
После объединения хромосомных наборов обоих пронуклеусов без всякого перерыва начинается митотическое деление ядра зиготы. За этим первым делением - следует серия следующих делений ядер и цитоплазмы, общие свойства которых таковы: 1. Разделившиеся клетки зародыша не растут, т. е. в промежутке между делениями масса их цитоплазмы не увеличивается. В результате суммарный объем и масса всех возникших клеток не превышает объема и массы яйцеклетки во время оплодотворения; 2. Между тем количество ДНК в ядре удваивается после каждого деления, как и при обычном митозе, так что все клетки сохраняют диплоидность. Эта серия делений называется дроблением яйцеклетки. Действительно, из-за отсутствия роста клеток после деления яйцеклетка как бы дробится на все более мелкие клетки. Последние называются бластомерами, а разделяющие их плоскости - бороздами дробления. Таким образом, дробление - это многократные митотические деления зиготы, в результате которых зародыш становится многоклеточным, не меняя при этом существенно своего объема.
Образование многоклеточности - первая и основная биологическая функция дробления. Вторая его функция состоит в увеличении так называемого ядерно-плазменного отношения. Еще задолго до возникновения современных представлений о роли ДНК в клеточном метаболизме было понято, что для нормальной жизнедеятельности клетки должно поддерживаться определенное отношение между количеством ядерного и цитоплазматического вещества. Это отношение было названо ядерно-плазменным отношением и обозначалось как я/пл.
Основные способы гаструляции
После того как зародыш достиг стадии бластулы, в нем начинаются интенсивные передвижения отдельных клеток и обширных участков стенки бластулы, приводящие, в конце концов, к тому, что более или менее однородный перед этим зародыш расчленяется на два или три слоя, которые называются зародышевыми листками. Самый внутренний зародышевый листок называется энтодермой, внешний - эктодермой. На эти листки расчленяются зародыши всех многоклеточных животных: лишь у губок дальнейшая судьба листков настолько необычна, что некоторые авторы избегают говорить применительно к ним об экто- и энтодерме. У всех животных, кроме губок и кишечнополостных, формируется средний зародышевый листок - мезодерма, располагающийся между двумя первыми. Процесс расчленения зародыша на зародышевые листки называется гаструляцией, а сам зародыш на стадии расчленения - гаструлой.
Способы гаструляции довольно разнообразны. Они отчасти связаны со строением бластулы, но эта связь далеко не однозначна. Особенно разнообразны типы гаструляции у низших беспозвоночных - кишечнополостных. У них распространен иммиграционный тип гаструляции, который был открыт в 1886 г. И. И. Мечниковым у некоторых гидромедуз и может считаться эволюционно наиболее древним. Этот процесс сводится к вселению в полость бластоцеля отдельных клеток, выклинивающихся из стенки бластулы. Иногда процессы иммиграции идут без особого порядка по всей поверхности бластулы. Тогда говорят о мультиполярной иммиграции. Большей же частью выселение идет с одного определенного полюса- униполярная иммиграция.
Известна также биполярная иммиграции, когда выселение идет с противоположных полюсов.
У тех кишечнополостных, где дробление заканчивается морулой без полости, наблюдается другой тип гаструляции, который получил название деламинация (расслоение). Он ограничивается выравниванием внутренних стенок клеток наружного слоя, причем такое выравнивание нередко идет волной, от одной соседней клетки к другой. Вдоль выровненных поверхностей формируется базальная мембрана, отделяющая этот внешний клеточный слой (эктодерму) от внутренней массы клеток, которая вся становится энтодермой. При деламинации, таким образом, клеточные перемещения почти отсутствуют.
Наконец, некоторым высшим кишечнополостным (сцифоидные медузы, коралловые полипы) присущ иной тип гаструляции, широко распространенный у более высших форм: впячивание, или инвагинация (впрочем, небольшое впячивание в месте униполярной иммиграции обнаружено и у некоторых гидроидных полипов). В этих случаях внутрь бластоцеля входят не отдельные клетки, а клеточный пласт, не утративший эпителиальной структуры. Впрочем, этот способ гаструляции легко заменяется другим, более примитивным. Так, сцифоме-дузе Aurelia flavldula свойственна более или менее выраженная инвагинация, Aurelia marginalis - мультиполярная иммиграция, Aurelia aurita - нечто вроде униполярной иммиграции с последующей эпитализацией. Для ряда видов гидроидных полипов тоже характерны различные сочетания иммиграционных и деламинационных процессов или же у них последовательно протекают оба процесса. Во всяком случае, гаструляционные процессы у кишечнополостных крайне вариабельны.
В других группах животных деламинация и иммиграция тоже представляют собой компоненты гаструляционного процесса. Например, у иглокожих путем иммиграции с вегетативного полюса закладывается так называемая первичная мезенхима, из которой потом формируются некоторые временные органы личинки (скелет, органы выделения). В целом же процесс гаструляции приобретает более организованный характер и осуществляется обычно путем инвагинации вегетативной стенки бластулы. Полость вворачивания называется гастроцелем, а ведущее в нее отверстие - бластопором (первичным ртом). Края бластопора называются его губами.
Так как при инвагинации механическая целостность стенки бластулы не нарушается, очевидно, что вворачивание дна бластулы должно сопровождаться более или менее значительным смещением клеточного материала боковых стенок в вегетативном направлении (вегетопетально). Действительно, такие движения всегда происходят, и скорость их, как правило, не меньше скорости вворачивания. Вегетопетальные движения того слоя, который в данный момент еще находился па поверхности гаструлы, называют эпиболией (обрастанием). Существует немало случаев чисто эпиболической гаструляции, когда инвагинация невозможна из-за малых размеров бластоцеля. или инертности крупных, богатых желтком вегетативных макромеров. Так обстоит дело, например, у ряда малощетинковых червем: макромеры здесь просто накрываются наползающими на них микромерами.
Материал, оставшийся на поверхности зародыша после завершения гаструляции, есть наружный зародышевый листок, или эктодерма. Что касается погрузившегося любым способом внутрь материала, то лишь у кишечнополостных он представляет собой чистую энтодерму - внутренний зародышевый листок, формирующий впоследствии стенку пищеварительного тракта с его производными. У всех вышестоящих систематических групп погрузившийся внутрь в ходе гаструляции материал содержит кроме энтодермы еще и материал будущего среднего зародышевого листка - мезодермы, который потом отделяется от энтодермы.
Гаструляции у амфибий
Гаструляция амфибий - сложный комплексный процесс, состоящий из множества разнородных клеточных движений. Основными его компонентами принято считать эпиболию и инвагинацию. В первом приближении это можно принять, но не следует забывать, что сами названные процессы имеют составной характер, и, кроме того, они дополняются процессами иммиграции и деламинации. Как мы знаем, вегетативная стенка бластулы амфибий сложена из крупных, богатых желтком макромеров. Поэтому на вегетативном полюсе не может возникнуть такое обширное впячивание, как у иглокожих и ланцетника. Однако, по-видимому, некоторые богатые желтком наружные макромеры все же погружаются внутрь зародыша.
Эти движения иммиграционного типа получили название предгаструляционных. Они приводят к сокращению светлой вегетативной зоны на поверхности зародыша и к соответственному увеличению темной (пигментированной) анимальной зоны. Последний процесс можно рассматривать как первую, пассивную фазу эпиболии.Собственно гаструляция начинается в области уже известного нам серого серпа. Там сначала возникает выровненная линия клеточных стенок, проходящая чуть вегетативнее границ анимального (пигментированного) и вегетативного (светлого) полушарий, а затем по этой линии образуется узкая, идущая вглубь щель - зачаток бластопора. Щелевидное впячивание углубляется, вовлекает в себя все новые клетки поверхности зародыша и принимает вид серповидной бороздки. Анимальный верхний край этой бороздки называется спинной, или дорсальной, губой бластопора, так как здесь расположен задний край спинной стороны зародыша. Полость щелевидной бороздки несколько расширяется и превращается в зачаток первичной кишки, или архентерона.
Дальнейший ход гаструляции связан, прежде всего, с подворачиванием клеточного материала через дорсальную губу бластопора: клетки анимальных районов смещаются в вегетативном направлении (вегетопетально) вплоть до губы бластопора и, подвернувшись через нее, образуют дорсальную выстилку углубляющегося архентерона. Таким образом, из вышесказанного видно, что клеточный состав дорсальной губы бластопора непрерывно обновляется.
Вегетопетальные движения клеток наружной поверхности гаструлы в направлении дорсальной губы бластопора представляют собой продолжение движений эпиболии. В результате этих движений бластопор смещается в вегетативном направлении, и площадь поверхности, занимаемая анимальными клетками, все время увеличивается.
Движения эпиболии осуществляются главным образом благодаря следующим двум процессам: 1. Активному схождению (конвергенции) к средней (сагиттальной) линии зародыша клеток так называемой супрабластопоральной области, расположенной непосредственно дорсальнее бластопора. Схождение клеток сопровождается их переупаковкой - сменой соседей. Из-за этого схождения данный участок сужается в поперечном и растягивается в продольном направлении. На зародышах шпорцевой лягушки измерена скорость сдвига, являющегося следствием данного растяжения: она составляет примерно 3,5 мкм/мин. 2. Вдвижению друг между другом клеток внутренних слоев крыши бластоцеля в более анимальных, удаленных от бластопора районах зародыша. В результате такого вдвижения крыша бластоцеля растягивается примерно равномерно по всем направлениям. Оба движения осуществляются согласованно и способствуют увеличению площади анимальной части зародыша, т. е. эпиболии.
Тем временем бластомер продолжает расти в стороны, охватывая, светлую вегетативную зону сначала полукольцом, а потом и полным кольцом, которое по ходу гаструляции постепенно уменьшается до узкого отверстия. Заключенный внутри кольцевидного бластопора светлый вегетативный материал называется желточной пробкой. В кольцевидном бластопоре кроме известной уже нам дорсальной губы различают вентральную дубу (участок, противоположный дорсальной губе) и боковые губы. Через эти губы тоже идет подворачивание материала, но оно несравненно слабее, чем подворачивание через дорсальную губу.
Ввернувшийся внутрь клеточный материал стенки архентерона продвигается сплошным слоем по внутренней поверхности стенки бластоцеля, постепенно оттесняя бластоцель в вентральном направлении и, в конце концов, вытесняя его почти полностью. Это движение называют инвагинацией, но, как и движение эпиболии, оно слагается из ряда компонентов.
Самая ранняя стадия инвагинации (закладка бластопора) связана с возникновением в области бластопора группы так называемых колбовидных клеток с узкими апикальными «шейками» и удлиненными, раздутыми телами. Впячивание бластопора закладывается как раз благодаря активному сужению «шеек» этих клеток и растяжению клеточных тел. Структурные основы этого и других морфогенетических процессов рассматриваются в конце главы. При дальнейшей инвагинации решающее значение имеют следующие процессы.
1. Клетки вершины гастрального впячивания (в том числе бывшие колбовидные), выбрасывая длинные отростки, активно ползут по стенке бластоцеля. Внутренняя выстилка этой стенки к данной стадии развития видоизменяется так, чтобы способствовать направленной миграции по ней клеток: здесь формируются волокна внеклеточного матрикса, состоящие из белка фибронектина и ориентированные в передне-заднем направлении зародыша, т. е. как раз вдоль траекторий движения клеток. На завершающей стадии инвагинации бывшие колбовидные клетки уплощаются и формируют выстилку передней части эмбрионального кишечника. Впоследствии они входят в закладку печени.
2. Клетки внутренних слоев дорсальной губы бластопора при подворачивании через губу резко меняют свою структуру и характер контактов: до подворачивания они представляют собой довольно плотно скрепленные столбчатые клетки эпителиального характера, а в процессе подворачивания превращаются в округлые разобщенные клетки, почти не контактирующие друг с другом. Таким образом, клеточные слои при подворачивании как бы рассыпаются на отдельные клетки. Такое рассыпание считается одним из основных факторов, способствующих подворачиванию. Впоследствии контакты между ввернувшимися клетками вновь восстанавливаются, но судьба ввернувшихся клеток уже необратимо отличается от судьбы клеток, оставшихся в супрабластопоральной области.
3. По мере вворачивания дорсальная губа бластопора смещается в вегетативном направлении. Такое смещение связано с тем, что скорость эпиболии (как уже говорилось выше, около 3,5 мкм/мин) существенно превышает скорость подворачивания клеточного материала через дорсальную губу (около 2,5 мкм/мин). В том же направлении, т. е. к вегетативному полюсу удлиняется деламинациояная борозда, разграничивающая ввернувшийся и еще не ввернувшийся клеточный материал. Удлинение деламинационной борозды - неотъемлемый компонент гаструляции амфибий.
Карты презумптивных зачатков зародышей амфибий
Какое положение займут различные районы бластулы после завершения гаструляции, и какова их окончательная судьба? Это можно установить, нанося на поверхность бластулы метки красками или другими веществами и прослеживая движение меток в ходе гаструляции. Результаты исследования выражают, отмечая на схеме бластулы или ранней гаструлы судьбу каждой меченой точки. Эти схемы названы картами презумптивных (будущих, или, в более точном переводе с латинского, предполагаемых) зачатков. Первым составил такие карты для зародышей амфибий немецкий эмбриолог В. Фохт (Vogt) в 20-х гг. нашего столетия. Он пропитывал кусочки агар-агара красящими веществами, которые поглощались живыми тканями и были для них безвредными (так называемые витальные краски - нильский голубой, нейтральный красный и др.), и прижимал эти кусочки к разным местам поверхности бластулы. Краска диффундировала в зародыш, и определенный его участок прокрашивался. Прослеживая перемещения окрашенного участка, можно было точно судить о том, куда он попадает в ходе гаструляции и в какой зачаток превратится. Позднее методы прижизненного мечения были усовершенствованы, и в составленные Фохтом карты внесены некоторые уточнения и исправления, касающиеся главным образом локализации презумптивной мезодермы. Мы изложим сначала классические данные Фохта, а затем упомянем о новейших поправках.
По Фохту, перед началом гаструляции все закладки зародыша расположены на поверхности, точнее - выходят на поверхность. Анимальнее щелевидной бороздки бластопора располагается зачаток так называемой прехордальной пластинки (прехорды), из которой по завершении гаструляции развивается главным образом выстилка ротовой полости.
Анимальнее прехордальной пластинки располагается зачаток будущей хорды. Дорсоанимальную часть зародыша занимает презумптивная эктодерма нервной системы (нейроэктодерма), а вентроанимальную часть-эктодерма покровов тела. Две последние закладки и по завершении гаструляции остаются на поверхности тела зародыша. Вегетативнее их располагается последовательно материал осевой мезодермы (идущей на образование туловищных и хвостовых сомитов), боковой пластинки (несегментированная часть мезодермы) и, наконец, энтодермы. Прехордальная пластинка, хорда, мезодерма (осевая и несегментированная) и энтодерма в ходе гаструляции погружаются внутрь зародыша. При этом первые две закладки подворачиваются через дорсальную губу, мезодерма - через боковые и вентральную губы, а энтодерма накрывается сходящимися губами бластопора. По изложенным данным, материал хорды и мезодермы должен после своего вворачивания непосредственно выстилать полость архентерона, образуя ее дорсальную стенку. В таком случае строение стенки архентерона у зародышей амфибий было бы подобным (гомологичным) строению этой же стенки у зародышей ланцетника или (за вычетом хорды) у зародышей иглокожих.В последующие годы было выявлено, что применительно к амфибиям такие выводы справедливы лишь для отряда Хвостатых (Urodela). Что касается бесхвостых амфибий (Anura), то, как было установлено Т. А. Детлаф, С. Левтрупом и Р. Келлером, у них материал хорды, осевой мезодермы и боковой пластинки ни на одной стадии развития не выходит на поверхность зародыша, а локализован с самого начала во внутренних слоях его стенки. При гаструляции этот материал подворачивается аналогично наружному, но никогда не приходит в контакт с гастральной полостью. Последняя оказывается выстланной с вентральной стороны крупными клетками желточной энтодермы, а с дорсальной стороны тонким слоем клеток, отделяющих эту полость от хорды и сомитов. Этот слой клеток принято называть гипохордой.
С учетом этой поправки классические карты презумптивных зачатков можно считать справедливыми для зародышей всех классов амфибий.
Современные методы маркировки эмбриональных тканей позволяют получать карты презумптивных зачатков не только для стадии бластулы, но и для более ранних стадий развития, в частности для периода дробления. Такая маркировка осуществляется путем инъекции в отдельные бластомеры флуоресцентных красителей, которые затем выявляются в потомках инъецированного бластомера при рассмотрении гистологических срезов в люминесцентном микроскопе.
Использование такого метода позволило прийти к выводу принципиальной важности: к стадии 32 бластомеров презумптивное значение большинства бластомеров еще не определено с полной точностью - один и тот же бластомер может в различных процентах случаев давать разные закладки и, напротив, разные бластомеры - одни и те же закладки. Например, сомиты могут образовываться из тех же бластомеров, что хорда и нервная трубка (бластомер В1), или из тех же бластомеров, что и боковая пластинка (бластомеры ВЗ, СЗ, В4, С4). С другой стороны, нервная трубка может возникать из любого следующего бластомера: А1, А2, В1, В2, ВЗ, С1 или С2, хотя в других случаях из двух последних бластомеров может возникнуть энтодерма. Это связано с тем, что морфогенетические движения клеток в последующем развитии не идеально точны: потомки какого-либо одного бластомера могут до некоторой степени случайно перемешиваться с потомками другого бластомера. Но поскольку это не приводит к нарушениям структуры организма, очевидно, что по крайней мере в период дробления и до его начала (в период ооплазматической сегрегации) судьба бластомера окончательно не определяется. Даже после завершения гаструляции возможно взаимное перемешивание, а значит, и переопределение судеб отдельных клеток.
Надо особо подчеркнуть, что карты презумптивных зачатков, какими бы методами они не составлялись, дают сведения о судьбе отдельных участков зародыша лишь в его нормальном развитии и ничего не говорят о том, может или нет судьба клеток быть переопределена при их перемещении в другое положение. Иными словами, карты не дают сведений о степени детерминации клеточной судьбы. Подобного рода данные рассматриваются в следующей главе.
Нейруляция и формирование осевых органов у зародышей амфибий
Движения гаструляции у зародышей позвоночных без существенного перерыва переходят в движения, связанные с нейруляцией - закладкой центральной нервной системы. Нейруляция - характернейший для всех позвоночных формообразовательный процесс, определяющий главные структурные особенности представителей этого типа. Зародыш позвоночных в период нейруляции называется нейрулой. Мы рассмотрим процесс нейруляции на примере амфибий.
Обычно нейруляцию определяют как процесс скручивания нейральной эктодермы, расположенной на спинной стороне зародыша, в нервную трубку. В действительности это лишь часть тех формообразовательных движений, которые происходят в зародыше по окончании гаструляции. В целом эти движения состоят в конвергентном (сходящемся) смещении материала эктодермы и мезодермы к средней линии спинной стороны зародыша (вентродорсальные движения); происходит также растяжение дорсальной эктодермы зародыша в передне-заднем направлении.
Собственно нейруляционные движения в презумптивной нейральной эктодерме представляют собой часть этих движений и развиваются на их основе. Сначала нейральная эктодерма уплощается и превращается в нервную пластинку, которая в головной части зародыша шире, чем в туловищной. Края пластинки приподнимаются и образуют нервные валики, окаймляющие пластинку сплошной подковой. Затем поверхность нервной пластинки начинает довольно быстро сокращаться в поперечном направлении преимущественно за счет погружения ее наружных клеток в ее же внутренние слои. Одновременно она начинает складываться по средней линии. Возникающее по средней линии углубление нервной пластинки называется нервным желобом. Еще чуть позже края нервной пластинки смыкаются, и образуется нервная трубка, полость внутри которой называют невроцелем. Передняя расширенная часть нервной трубки превращается в головной мозг, а ее невроцель - в полость мозгового пузыря. Более узкая туловищная часть трубки превращается в спинной мозг, а его полость - в спинномозговой канал.
После смыкания нервного желобка в нервную трубку материал нервных валиков, расположенный сначала по периферии нервной пластинки, концентрируется вдоль средней линии зародыша дорсальнее трубки в виде образования, напоминающего петушиный гребень. Поэтому данную структуру называют нервным гребнем. Клетки нервного гребня не входят в состав центральной нервной системы; они дают множество различных производных.
Еще до начала скручивания нервной пластинки в трубку или в самом начале этого скручивания из осевой мезодермы точно по средней (сагиттальной) линии зародыша в виде тяжа обособляется хорда, или спинная струна. Хорда существует длительное время, вплоть до формирования скелетных позвонков, которыми она почти нацело вытесняется. Располагается хорда под туловищной частью нервной трубки; ее передний конец как раз совпадает с границей туловищного и головного отделов. Спереди от хорды находится тонкий пласт клеток прехордальной пластинки, образующих выстилку глотки и ротовой полости.
Сразу латеральнее материала хорды располагается мезодерма будущих сомитов; вентральнее границы нервной пластинки и покровной эктодермы этот материал плавно переходит в мезодерму боковой пластинки. Внутри зачатков сомитов возникает полость, переходящая в узкую щель, разделяющую боковую пластинку на два листка: париетальный, прилежащий к покровной эктодерме, и висцеральный, прилежащий к энтодерме. Внутренняя полость и щель образуют уже знакомую нам вторичную полость тела - целом. У зародышей амфибий, как и у подавляющего большинства других позвоночных животных, целом возникает путем расхождения клеток, т. е. шизоцельным путем. Только применительно к ланцетнику и некоторым акуловым рыбам можно говорить об энтероцельной его закладке (т. е. об отшнуровке от единой полости архентерона).
Формирование хорды и сомитов связано с интенсивной миграцией материала эмбриональной мезодермы в вентродорсальном направлении, к средней линии зародыша, т. е. к линии образования хорды. Таким образом, хорда образуется на месте встречи двух клеточных потоков, причем встречно движущиеся клетки втискиваются друг между другом. Это явление названо интеркаляцией клеток.
В результате интеркаляции хорда удлиняется. Вентродорсальное движение мезодермы (совпадающее по направлению с нейруляционными движениями) характерно для туловищной области зародыша. В шейной и головной областях происходит обратное, дорсовентральное движение мезодермальных клеток, концентрирующихся на вентральной стороне тела, в области будущей закладки сердца.Вскоре после обособления хорды, еще до завершения нейруляции начинается метамеризация осевой мезодермы, т. е. ее разделение на парные сегменты - сомиты. Это один из важнейших морфогенетических процессов у позвоночных, закладывающий основы их опорно-двигательной системы. Метамеризация мезодермы идет в направлении спереди назад. У амфибий она продолжается и после вылупления зародыша из яйцевых оболочек, по мере роста ее хвоста, где один за другим формируются хвостовые сомиты (из материала заднего отдела нервной трубки). Клеточные механизмы метамеризации у разных позвоночных различны. У бесхвостых амфибий в процессе метамеризации клетки осевой мезодермы поворачивается на 90°, изменяя первоначальную поперечную ориентацию на продольную.
У хвостатых амфибий формирование сомитов связано с группированием мезодермальных клеток в своеобразные «розетки», у зародышей птиц - в сходные с розетками веерообразные структуры, постепенно достраивающиеся до полного сомита.
Механизмы морфогенетических движений гаструляции и нейруляции
Несмотря на все разнообразие описанных выше гаструляционных и нейруляционных морфогенетических движений, все они, равно как и морфогенетические движения, в последующем развитии, основаны на немногих клеточных и молекулярных процессах, а также регуляторных механизмах. Рассмотрим их более подробно.
Активность морфогенетических движений. Прежде всего, надо отметить следующее: подавляющее большинство морфогенетических движений на протяжении всего развития являются активными; это значит, что их источники энергии и исполнительные механизмы находятся внутри клеток как раз того участка зародыша, который испытывает данную деформацию. Это обобщение выработалось в эмбриологии не сразу: на заре исследования морфогенетических движений многие авторитетные исследователи считали, что, например, гаструляционное и нейруляционное впячивания возникают пассивно, в результате бокового давления на впячивающийся клеточный материал. Такое боковое давление могло возникать, по их мнению, за счет размножения клеток сбоку от впячиваний: размножающиеся клетки как бы сминают расположенный между ними участок пласта, заставляя его впячиваться.
Однако эта точка зрения в подавляющем большинстве случаев не получила экспериментального подтверждения. Лучшим свидетельством против нее являются многочисленные эксперименты по изоляции участка, которому предстоит впячиваться, от боковых областей, от которых должно было исходить давление: при такой изоляции впячивание не только осуществляется, но даже протекает быстрее. Отсюда следует, что боковые участки зародыша не только не способствуют впячиванию, но, напротив, оказывают ему сопротивление ввиду своего растяжения. О факторе растяжения тканей и его роли в регуляции морфогенетических движений мы еще поговорим позже.
Поляризация клеток. Любое активное изменение формы эпителиального пласта в период гаструляции и нейруляции, а также в последующих органогенезах, начинается с того, что клетки данного участка пласта поляризуются, то есть вытягиваются в направлении, перпендикулярном или косом к поверхности пласта.
Поляризация клеток эмбриональных эпителиев - пример согласованного коллективного клеточного поведения. Клетки эмбриональных эпителиев практически никогда не поляризуются поодиночке, а всегда целыми группами. Нередко можно проследить волну поляризации, распространяющуюся от одной клетки к другой. Такая волна наблюдается, например, при подворачивании клеток через дорсальную губу бластопора: у зародышей шпорцевой лягушки в процессе подворачивания каждая последующая клетка поляризуется, удлиняясь более чем вдвое, причем каждая последующая клетка поляризуется в течение 3-5 мин. Таким образом, за час поляризуется и подворачивается около 20 клеток, что соответствует инвагинации участка пласта поперечником примерно 200-300 мк. Интенсивная поляризация клеток происходит и в нейроэктодерме при формировании нервной пластинки. Таким образом формируется столбчатый нейроэпителий.
Поляризация клеток основана на сложных и еще недостаточно изученных перестройках цитоскелета и клеточной мембраны: сборке микротрубочек и микрофиламентов и их ориентации по длинной оси поляризующейся клетки, а также движениями так называемых интегральных (встроенных в мембрану) белков в плоскости плазматической мембраны. В результате этих движений перераспределяются ионные каналы и насосы: первые концентрируются преимущественно на апикальной (внешней) стороне поляризующихся клеток, вторые - на боковых и базальных сторонах. В поляризующихся клетках формируется также система связывающих их межклеточных контактов. Поляризация клеток обусловливает протекание в них направленного эндо- и экзоцитоза, о чем будет сказано позже.
Сокращение поляризованных клеток. Формирование сомкнутых зон (или, как иногда говорят, доменов) поляризованных клеток лишь указывает места будущих впячиваннй или выпячиваний, но не приводит непосредственно к изменению формы клеточного пласта. Форма пласта преобразуется в результате последующих изменений формы поляризованных клеток, выражающихся в сокращении определенных участков их поверхности или же всей поверхности. Один из простейших и широко распространенных процессов такого рода - сокращение апикальных поверхностей поляризованных клеток. Именно оно и приводит, в частности, к описанному выше сужению «шеек» колбовидных клеток. Аналогичное сокращение апикальных поверхностей клеток нейроэктодермы играет важную, хотя и не единственную, роль в скручивании нервной трубки. Сокращение апикальных поверхностей обусловлено, прежде всего «самопоеданием» клетками своей апикальной мембраны путем эндоцитоза (захватывания внутрь клетки мембранных пузырьков). По мнению ряда авторов, эти пузырьки направленно транспортируются в противоположные, базальные районы клетки и там встраиваются путем экзоцитоза, наращивая базальные отделы клеточной мембраны за счет апикальных.
Обычно сокращение не ограничивается апикальными клеточными поверхностями: боковые поверхности поляризованных клеток тоже сокращаются, выгибая при этом клеточный пласт. Это особенно ясно проявляется на примере веерообразных группировок из растянуто-скошенных клеток, образование которых предшествует любому впячиванию клеточного пласта. Еще в 1914 г. А. Г. Гурвич обнаружил, что в нейральных пластах зародышей позвоночных направление наклонов осей скошенных клеток как бы предсказывает еще до начала впячивания пласта, куда последний изогнется: изогнутая поверхность будет перпендикулярной осям скошенных клеток. Это правило «прогностических наклонов» клеточных осей объясняется тем, что изгиб пласта осуществляется именно за счет спрямления клеток - перехода формы их сечений от скошенной к прямоугольной. Но такое спрямление и является результатом сокращения боковых поверхностей клеток: понятно, что при неизменном объеме площадь поверхности прямоугольной клетки меньше поверхности скошенной клетки.
Сокращение боковых клеточных поверхностей - тоже активный процесс, в котором, вероятно, участвуют актиновые микрофиламенты.
Роль механических напряжений в организации гаструляционных и нейруляционных движений
Мы уже могли убедиться, что в результате гаструляционных и неируляционных движении возникает сложная и вместе с тем высокоупорядоченная, геометрически правильная организация зародыша. Благодаря чему устанавливается и поддерживается эта правильность? Почему многочисленные отдельные клетки и целые клеточные пласты движутся в строго определенных направлениях, образуя закономерно локализованные скопления клеток или изгибы пластов? Может быть, в каждую отдельно взятую клетку зародыша заранее «вложена» информация о се движении и пункте ее назначения? Такое предположение можно сразу же отклонить, хотя бы ввиду описанного выше «статистического» характера карт презумптивных зачатков: мы уже видели, что даже в ходе ненарушенного, нормального развития движение и окончательная судьба участков зародыша не определяются с «поклеточной» точностью. Чисто случайно отдельные клетки могут двигаться разными путями и входить в состав различных зачатков без того, чтобы это нарушало правильную структуру целого организма. С другой стороны, если изолировать и заставить развиваться вне зародыша небольшие кусочки эмбриональных тканей размером в несколько сотен клеток, то, хотя из данного кусочка и будет, как правило, развиваться набор зачатков, соответствующий его презумптивной судьбе, форма и расположение этих зачатков будут неправильными. Эти данные наряду со многими другими говорят о том, что факторы организации движений гаструляции и нейруляции как-то связаны с целым зародышем. Что могут представлять собой эти организующие факторы?
Ряд данных показывает, что такими факторами могут являться механические натяжения тканей зародыша. Первым по времени фактором, обусловившим натяжение эмбриональных тканей, является тургорное давление в полости бластоцеля, растягивающее крышу бластоцеля. Это натяжение еще сравнительно незначительно. Однако оно заметно усиливается в ходе гаструляции, поскольку движения инволюции через губы бластопора (главным образом через дорсальную губу) приводят к растяжению всей поверхности зародыша в продольном (передне-заднем) направлении. Кроме того, инвагинирующий материал хордомезодермы, ползущий по выстилке бластоцеля, растягивает как сам себя, так и расположенный перед ним участок стенки бластоцеля, на который он наползает.
Роль эпителиально-мезенхимных взаимодействий в дифференцировке энтодермальных зачатков. Для дифференцировки энтодермальных зачатков требуются непосредственные контакты с мезодермой, причем на ранних стадиях развития менее специфические, а для окончательной дифференцировки - более специфические контакты. Так, для формирования выроста легкого из эпителия передней кишки достаточен контакт эпителия с мезенхимой этого же зачатка. Добавление чужеродной мезенхимы может полностью изменить направление развития зачатка: под влиянием мезодермы желудка легочная энтодерма будет образовывать структуры, сходные с железами желудка, под влиянием мезодермы печени - печеночные тяжи. Для начальных стадий морфогенеза зачатка печени необходим его контакт с мезодермальными клетками зачатка сердца, а для дальнейшей биохимической дифференцировки клеток печени - контакт с собственной, печеночной мезодермой. Присутствие специфической мезодермы необходимо также для полной дифференцировки и функционирования щитовидной железы. Несколько менее специфические влияния требуются при развитии поджелудочной железы: для нормальной дифференцировки эпителия поджелудочной железы в клетки, секретирующие гормоны (в том числе инсулин), также необходим контакт с мезенхимой, но в условиях эксперимента собственная мезенхима поджелудочной железы может быть заменена чужеродной мезенхимой слюнных желез или вторичной почки.
Гаструляция и нотогенез у человека протекают более 2 недель и приводят к образованию первичных эмбриональных органов - зародышевых листков и осевых органов (хорда, нервная трубка). Они детерминируют (предопределяют) общий план строения человеческого организма, служат источником развития вторичных органов, провизорных (существуют только у зародыша) и дефинитивных.
Название гаструляции дал английский ученый Э. Геккель (гастрея, лат. - выпуклость или чрево сосуда): наиболее примитивная, инвагинационная форма гаструлы напоминает сосуд, как и губки, представители кишечнополостных, наиболее примитивных дифференцированных многоклеточных животных.
Гаструляция начинается в процессе нидации (внедрения) бластоцисты в толщу эндометрия (иначе - имплантация, с лат. - сажать в): выделяя ферменты, зародыш разрушает оолемму и прилежащие клетки эндометрия в 6,5-7,5 сутки беременности. Одновременно эмбриобласт расщепляется (деламинация) на 2 слоя: наружный слой или зачаток, эпибласт - источник развития эктодермы (наружного зародышевого листка), нейроэктодермы, хорды и мезо дермы (среднего зародышевого листка); внутренний слой или зачаток, гипобласт - источник развития кишечной и желточной энтодермы. Отделение гипобласта начинается в каудальном (хвостовом) отделе эмбриобласта.
Разрастаясь и расслаиваясь, эпибласт образует амниотический пузырек, а гипобласт - желточный пузырек. Участок их соединения определяется как двухслойная гаструла. На 11-е сутки эмбриогенеза она ясно выражена, имеет форму овального диска (зародышевый щиток). В 2 недели его средний диаметр равен 0,2 мм, а бластулы в целом (плодного яйца) - 2,5 мм.
Каудальный край зародышевого щитка обращен к зародышевому стволу-месту сужающегося перехода эмбриобласта в трофобласт («амниотическая ножка»). На 14-е сутки эмбриогенеза именно каудальный конец гаструлы обнаруживает повышенную морфогенетическую активность: клетки эпибласта пролиферируют (увеличиваются в количестве), выселяются из него иммиграция и образуют строго по средней линии скопление - первичная полоска. Она содержит искривление в виде бороздки и удлиняется к будущему краниальном; (головному) концу зародышевого щитка. На переднем конце первичной полоски появляется утолщение - головной узелок Гензена. Первичная полоска детерминирует хвостовой отдел и двустороннюю симметрию тела человека характерную для всех позвоночных животных (у многих беспозвоночных теле построено по плану многолучевой симметрии). Таким образом, уже в конце 2-й недели эмбриогенеза можно определить дорсальную и вентральную (спинную и брюшную), правую и левую стороны, головной и хвостовой концы зародыша.
На третьей неделе эмбриогенеза формируется трехслойная гаструла (хордула или нейрула: хорда- струна, невро -жила/греч.). На 16-е сутки из первичной полоски, в обе стороны от нее выселяются клетки. Они образуют боковые пластинки («крылья») мезодермы. Мезодермальные пластинки проникают в промежутки между экто- и энтодермой, расщепляются и образуют 2 мезодермальных мешка. Таким образом определяются все 3 зародышевых листка.
На 18-е сутки узелок Гензена образует головной отросток (нотохорд) первичной полоски. Он входит в промежуток между экто- и энтодермой, а затем погружается в толщу энтодермы. На 19-е сутки клетки головного отростка образуют плотный осевой тяж (хорду или спинную струну) и осевую (парахордальную) мезодерму, из которой развиваются сомиты.
Одновременно (18-20-е сутки) над формирующейся хордой появляется нервная пластинка. Она имеет вид продольного тяжа, который состоит из крупных, темных клеток нейроэктодермы. Они активно пролиферируют размножаются). Поэтому в плотном окружении нервная пластинка искривляется и погружается вглубь зародышевого щитка, образует нервный желобок. На 4-й неделе края нервного желобка смыкаются, он превращается в нервную трубку. Таким образом протекает развитие осевых органов - нотогенез (эмбрионы 2,5-3,5 неделе).
В конце 3-й неделе длина зародышевого щитка достигает 2 мм при ширине 75 мм, тогда как средний диаметр плодного яйца превышает 6 мм (в 20-25 раз крупнее бластоцисты).
В конце 3-й неделе начинается стадия эмбрионального органогенеза, процессе которого образуются закладки всех дефинитивных систем органов определяется в общих чертах план дефинитивного строения человека. Уже конце 3-й недели обнаруживаются зачатки глаз, внутреннего уха, сердца, первые сомиты. Эмбриональный органогенез протекает в связи с начинающейся плацентацией.
Одновременно изменяется форма зародыша в результате неравномерного юта его частей: преобладает удлинение спинной части в срединной зоне (давление хорды и нервной трубки) и головного конца, где нервная трубка утолщается и образует зачаток головного мозга. Он значительно расширяется и огибает спереди хорду. В результате на 4-й неделе зародышевый щиток изгибается в поперечном направлении, сворачиваясь в трубку, и в переднее - заднем травлении (сближение головного и хвостового концов). Обособление тела зародыша продолжается до середины второго месяца и сопровождается разделением желточного пузырька на две части: дорсальная или зародышевая - первичная кишка, вентральная - желточный мешок. Первичная кишка является ровным источником развития пищеварительной и дыхательной систем, а желточный мешок - местом первичного кроветворения и образования кровеносных сосудов. Одновременно амниотический пузырек также разделяется на две части: зародышевая - кожа, внезародышевая - амнион (водная оболочка зародыша). Кроме того, обнаруживается неравномерный локальный рост тела зародыша. На 4-й неделе под увеличивающейся головой образуется сердечный горб: сердце достигает громадных относительных размеров, через неделю его угоняет», а затем и превышает печеночный горб. Их рост, а также увеличение пупочного стебелька в связи с формированием физиологической пупочной грыжи способствуют разгибанию и расхождению головы и хвоста у эмбрионов 6-7 неделе. На 7-й неделе хвост эмбриона значительно уменьшается, а голова достигает наибольших относительных размеров, как и живот, благодаря громадной печени. Дифференцируется, удлиняется и утоньшается шея с разгибанием головы, формируется лицо. В течение второго месяца вычленяются, удлиняются и разделяются на дефинитивные отделы конечности. На 8-й неделе начинается разделение пальцев руки, уменьшаются печень и живот. На десятой неделе утробной жизни редуцируется пупочный грыжевой мешок.
Четвертая неделя эмбриогенеза - это стадия наиболее интенсивного образования сомитов. Они определяют сегментарное или метамерное строение зародыша: его тело состоит из следующих друг за другом отрезков, сходных по строению - сегментов или метамеров. Метамеры дорсальной мезодермы называются сомитами (греч.: soma - тело). У эмбрионов 10 мм длины (5,5 неделе) их общее число достигает 43-44 пар. Признаки метамерного строения сохраняются в туловище человека на всю жизнь: 1) сегментарное строение собственного аппарата спинного мозга; 2) сегментарное строение позвоночного столба и связанных с ним глубоких собственных мышц спины; 3) сегментарное строение грудной клетки, включая межреберные мышцы, сосуды и нервы; 4) сегментарный выход спинномозговых нервов на протяжении спинного мозга; 5) метамерное размещение париетальных (пристеночных) ветвей нисходящей аорты.
Сомиты (первичные сегменты тела) лежат по обе стороны от хорды и разделяются на 3 части: 1) наружная (латеральная) - дерматом, источник развития соединительнотканной основы кожи; 2) внутренняя (вентромедиальная) - склеротом, источник развития скелета; 3) промежуточная (дорсомедиальная) - миотом, зачаток скелетной мускулатуры. Сомиты располагаются по обе стороны от нервной трубки и дорсальной аорты. Их ветви растут к сомитам и приобретают метамерное положение. Сомиты связаны со спланхнотомами сомитными ножками или нефротомами - суженная, промежуточная мезодерма, источник развития предпочки и первичной почки. Каудальный отдел промежуточной мезодермы не сегментируется и образует метанефрогенныи тяж, источник развития нефронов окончательной почки.
Сплахнотом (парная боковая пластинка мезодермы) разделяется на 2 листка: висцеральный (внутренностный) - спланхноплевра, окружает энтодерму туловищной кишки, участвует в развитии кишечной трубки и ее производных; париетальный (пристеночный)- соматоплевра, участвует в формировании стенок целомической полости зародыша и ее производных.
Мезодерма спланхнотомов служит источником развития мезотелия и главным источником мезенхимы, из которой развиваются все виды соединительной ткани, гладкая мышечная ткань, миокард. Мезенхима- это совокупность полиморфных (разных формы и строения) клеток разного происхождения и с различной судьбой. Первоначально мезенхимные клетки имеют звездчатую форму и благодаря своим длинным тонким отросткам образуют сеть с широкими интерстициальными (межуточными) каналами, по которым осуществляется дососудистая циркуляция тканевой жидкости. Вскоре после образования мезенхима теряет сетевидную структуру и уплотняется в результате увеличения концентрации клеток за счет митозов и продолжающегося выселения мезенхимных клеток из зародышевых листков и их производных.
Эктодерма служит источником развития кожного эпителия (эпидермиса), волос, ногтей; сальных, потовых и молочных желез; части эпителия ротовой полости и прямой кишки, моче- и семявыносящих путей; эмали зубов. Нейроэктодерма образует нейроны и особую соединительную ткань (нейроглию), а также нейрогипофиз и эпифиз, хроматофинные клетки.
Энтодерма является источником развития части выстилки воздухопроводящих путей и легочной паренхимы, части выстилки ротовой полости, эпителиев глотки, пищевода, желудочно-кишечного тракта, печени, поджелудочной железы, щитовидной и паращитовидных желез.
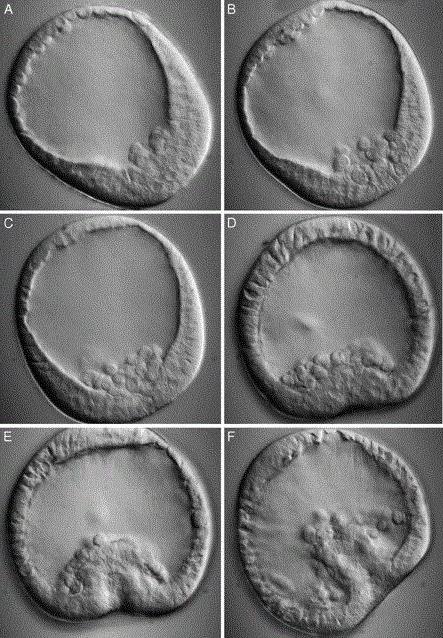
Рис. 7. Последовательные стадии (А - Г) гаструляции амфибий на сагиттальных разрезах:
А, А" - бластула; Б, Б" - ранняя гаструла; В, В" - средняя гаструла; Г, Г" - поздняя гаструла. Изображения А"- Г" повернуты на 90° относительно А-Г. блц - бластоцель; блп - бластопор; гц- гастроцель; д.г. - дорсальная губа бластопора; в.г. - вентральная губа бластопора; ж.пр.- желточная пробка {по Балинскому)

Рис. 8. Составные процессы гаструляции у амфибий:
А- схематическое строение ранней гаструлы, сагиттальный срез; Б-Г - схемы процессов, протекающих в соответственных областях гаструлы; Б - встраивание друг между другом клеток глубинных слоев эктодермы анимальной области (движения светлых клеток, показанные стрелками); В - краннокаудальное растяжение и поперечное сжатие супрабластопоральной области в результате переупаковки клеток (одинаковые клетки помечены одинаковыми номерами); Г - процессы подворачивания и «рассыпания» клеточного пласта на отдельные слабо связанные клетки в дорсальной губе бластопора; кк - колбовидные клетки; 1 - клетки до подворачивания; 2 - клетки после подворачивания (по Келлеру)

Рис. 9. Схема преобразования неполяризованной эмбриональной клетки у амфибий (А) в поляризованную (Б). Поляризация сопровождается перемещением клеточного ядра (я), желточных гранул (жг), элементов цитоскелета - микротрубочек (мт). Возникают новые межклеточные контакты (мк), часто ассоциированные с пучками микрофиламент (мф). Устанавливается направленный транспорт эндоцитозных пузырьков внутри клетки (стрелки)

Рис. 10. Последовательные стадии (А-Д) гаструляции у амфибий, внешний вид: б.г - боковые губы; в.г-вентральная губа; д.г - дорсальная губа бластопора; ж.пр. - желточная пробка (по Балннскому)

Рис. 11. Схема перемещений клеток, происходящих при гаструляции у амфибий (no: Waddington С.Н., из Карлсон, 1983):
А, Б, В - последовательные стадии перемещения клеток в ходе гаструляции: 7 - эктодерма, 2 - материал будущей мезодермы. 3 - богатые желтком клетки энтодермы, 4 - клетки в составе эктодермы, образующие презумлтивную нервную пластинку, 5 - бластопор, 6 - материал хорды, 7 - энтодерма, 8 - кожная эктодерма, 9 - бластоцель, 10 - гастроцель, или первичная кишка, 1; - материал нейроэктодермы, 12 - дорсальная губа бластопора, 13 - вентральная губа бластопора, 14 – материал мезодермы

Рис. 12. Схема образования первичной полоски и иммиграции клеток:
А - вид сверху на зародышевый щиток (стрелки - перемещение клеток в краниокаудальном направлении); Б - вид сбоку; 7 - гензеновский узелок, 2 - первичная ямка, 3 - первичная бороздка, 4 - материал мезодермы в составе первичной полоски, 5 - мезодерма, 6 - энтодерма, 7 - прехордальная пластинка, 8 - хордальный отросток (стрелки - направления перемещения клеток)
Сущность стадии гаструляции заключается в том, что однослойный зародыш - бластула - превращается в многослойный - двух- или трехслойный, называемый гаструлой (от греч. гастер - желудок в уменьшительном смысле).
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок -эктодерму -и внутренний зародышевый листок - энтодерму. Энтодерма формирует первичную кишку с полостью внутри-гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющими морфологическими признаками гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У позвоночных помимо двух упомянутых во время гаструляции образуется еще третий зародышевый листок - мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, представляющего собой хордомезодерму, является эволюционным усложнением фазы гаструляции у позвоночных и связано с ускорением у них развития на ранних стадиях эмбриогенеза. У более примитивных хордовых животных, таких, как ланцетник, хордомезодерма обычно образуется в начале следующей после гаструляции фазы - органогенезе. Смещение времени развития одних органов относительно других у потомков по сравнению с предковыми группами является проявлением гетерохронии. Изменение времени закладки важнейших органов в процессе эволюции встречается не редко.
Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий.
Способы гаструляции различны . Выделяют четыре разновидности направленных в пространстве перемещений клеток, приводящих к преобразованию зародыша из однослойного в многослойный.
Инвагинация - впячивание одного из участков бластодермы внутрь целым пластом. У ланцетника впячиваются клетки вегетативного полюса, у земноводных инвагинация происходит на границе между анимальным и вегетативным полюсами в области серого серпа. Процесс инвагинации возможен только в яйцах с небольшим или средним количеством желтка.
Эпиболия - обрастание мелкими клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных.
Деламинация -расслоение клеток бластодермы на два слоя, лежащих друг над другом. Деламинацию можно наблюдать в дискобластуле зародышей с частичным типом дробления, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деламинация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта.
Иммиграция - перемещение групп или отдельных клеток, не объединенных в единый пласт. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных.
В каждом конкретном случае эмбриогенеза, как правило, сочетаются несколько способов гаструляции.
Особенности стадии гаструляции.
Гаструляция характеризуется разнообразными клеточными процессами. Продолжается митотическое размножение клеток, причем оно имеет разную интенсивность в разных частях зародыша. Вместе с тем наиболее характерная черта гаструляции состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу. Происходит сортировка клеток по их принадлежности к разным зародышевым листкам, внутри которых они «узнают» друг друга.На фазу гаструляции приходится начало цитодифференцировки, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие овоплазматической сегрегации. Так, эктодермальные клетки земноводных имеют темный цвет из-за пигмента, попавшего в них из анимального полюса яйцеклетки, а клетки энтодермы - светлый, так как происходят из вегетативного полюса яйца.Во время гаструляции очень велика роль эмбриональной индукции. Показано, что появление первичной полоски у птиц - результат индукционного взаимодействия между гипобластом и эпибластом. Гипобласту присуща полярность. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски.Такие проявления целостности зародыша как детерминация, эмбриональная регуляция и интегрированность присущи ему во время гаструляции в той же мере как и во время дробления
30. Первичный органогенез (нейруляция) как процесс образования комплекса осевых органов хордовых. Дифференцировка зародышевых листков. Образование органов и тканей.
Органогенезы, заключающиеся в образовании отдельных органов, составляют основное содержание эмбрионального периода. Они продолжаются в личиночном и завершаются в ювенильном периоде. Органогенезы отличаются наиболее сложными и разнообразными морфогенетическими преобразованиями. Необходимой предпосылкой перехода к органогенезам является достижение зародышем стадии гаструлы, а именно формирование зародышевых листков. Занимая определенное положение друг по отношению к Другу, зародышевые листки, контактируя и взаимодействуя, обеспечивают такие взаимоотношения между различными клеточными группами, которые стимулируют их развитие в определенном направлении. Это так называемая эмбриональная индукция -важнейшее следствие взаимодействия между зародышевыми листками.
В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Постепенно развивается определенная форма органов, устанавливаются пространственные и функциональные связи между ними. Процессы морфогенеза сопровождаются дифференциацией тканей и клеток, а также избирательным и неравномерным ростом отдельных органов и частей организма. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель.
Самое начало органогенеза называют нейруляцией . Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку.Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры - сомиты.
Нервная система позвоночных, включая человека, отличается устойчивостью основного плана строения на протяжении всей эволюционной истории подтипа. В формировании нервной трубки у всех хордовых много общего. Вначале неспециализированная спинная эктодерма , отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку , представленную нейроэпителиальными клетками цилиндрической формы.
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри - невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитохалазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень . Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу и образуют два главных потока. Клетки одного из них-поверхностного -включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении , образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма , занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой . Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов - склеротомы -превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образуетмиотомы , из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы , которые дают начало внутреннему слою кожи - дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела - целом . Внутренний листок , прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов нервная трубка - хорда - кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляции.Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов. Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка. Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения; слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретирующие клетки передней и средней долей гипофиза, щитовидной и паращитовидной желез.
Мезодерма помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа.
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется и в итоге в его формировании принимают участие два или три зародышевых листка.
31. Провизорные органы хордовых. Группа Анамнии и Амниоты. Образование, строение, особенности функционирования и эволюции провизорных органов и зародышевых оболочек. Амнион, хорион или сероза, аллантоис, желточный мешок, плацента. Типы плаценты, ее значение.
У животных разных типов в период эмбрионального развития возникают провизорные эмбриональные органы , обеспечивающие жизненно важные функции: дыхание, питание, выделение, движение и др. Недоразвитые органы самого зародыша еще не способны функционировать по назначению, хотя обязательно играют какую-то роль в системе развивающегося целостного организма. Как только зародыш достигает необходимой степени зрелости, когда большинство органов способны выполнять жизненно важные функции, временные органы рассасываются или отбрасываются.
Время образования провизорных органов зависит от того, какие запасы питательных веществ были накоплены в яйцеклетке и в каких условиях среды происходит развитие зародыша. У бесхвостых земноводных, например, благодаря достаточному количеству желтка в яйцеклетке и тому, что развитие идет в воде, зародыш осуществляет газообмен и выделяет продукты диссимиляции непосредственно через оболочки яйца и достигает стадии головастика. На этой стадии образуются провизорные органы дыхания (жабры), пищеварения и движения, приспособленные к водному образу жизни. Перечисленные личиночные органы дают возможность головастику продолжить развитие. По достижении состояния морфофункциональной зрелости органов взрослого типа временные органы исчезают в процессе метаморфоза.
У пресмыкающихся и птиц запасов желтка в яйцеклетке больше, но развитие идет не в воде, а на суше. В связи с этим очень рано возникает потребность в обеспечении дыхания и выделения, а также в защите от высыхания. У них уже в раннем эмбриогенезе, почти параллельно с нейруляцией, начинается формирование провизорных органов, таких, как амнион, хорион и желточный мешок. Чуть позднее формируется аллантоис. У плацентарных млекопитающих эти же провизорные органы образуются еще раньше, поскольку в яйцеклетке очень мало желтка. Развитие таких животных происходит внутриутробно, образование провизорных органов у них совпадает по времени с периодом гаструляции.
Наличие или отсутствие амниона и других провизорных органов лежит в основе деления позвоночных на две группы: Amniota и Anamnia. Эволюционно более древние позвоночные, развивающиеся исключительно в водной среде и представленные такими классами, как Круглоротые, Рыбы и Земноводные, не нуждаются в дополнительных водных и других оболочках зародыша и составляют группу анамний. К группе амниот относят первичноназемных позвоночных, т.е. тех, у кого эмбриональное развитие протекает в наземных условиях.
Это три класса: Пресмыкающиеся, Птицы и Млекопитающие. Они являются высшими позвоночными, так как имеют скоординированные и высокоэффективные системы органов, обеспечивающие им существование в наиболее сложных условиях, каковыми являются условия суши. Эти классы насчитывают большое количество видов, вторично перешедших в водную среду. Таким образом, высшие позвоночные оказались в состоянии освоить все среды обитания. Подобное совершенство было бы невозможным, в том числе и.без внутреннего осеменения и специальных провизорных эмбриональных органов.
В строении и функциях провизорных органов различных амниот много общего. Характеризуя в самом общем виде провизорные органы зародышей высших позвоночных, называемые также зародышевыми оболочками, следует отметить, что все они развиваются из клеточного материала уже сформировавшихся зародышевых листков. Некоторые особенности имеются в развитии зародышевых оболочек плацентарных млекопитающих, о чем будет сказано ниже.
· Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
· Хорион (сероза) - самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция - дыхательный газообмен; у млекопитающих он выполняет гораздо более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, например гормонов.
· Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мездермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например в спланхноплевре желточного мешка, развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.У млекопитающих нет запасов желтка и сохранение желточного мешка может быть связано с важными вторичными функциями. Энтодерма желточного мешка служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке. Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
· Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя - сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
Плаценты различаются по форме и размещению ворсинок. По этому признаку выделяют следующие типы плацент . Диффузная – вся поверхность плодного пузыря равномерно покрыта ворсинками. Такая плацента характерна для свиньи. У жвачных животных наблюдается котиледонная плацента, где ворсинки собраны в группы – котиледоны. Поясная плацента характерна для хищных млекопитающих. В этом случае ворсинки окружают плодный пузырь в форме широкого пояса. Следующий тип плаценты – дискоидальная. Она наблюдается у обезьян и человека, когда ворсинки располагаются на плодном пузыре в виде диска.
Плацента имеет громадное значение для развивающегося детеныша.
Она выполняет ряд важных функций:
1) трофическую – посредством плаценты происходит питание плода;
2) дыхательную – осуществляет поступление кислорода;
3) экскреторную – происходит выделение в кровь материнского организма продуктов обмена;
4) защитную – предохраняет зародыш от проникновения различных бактерий;